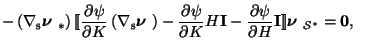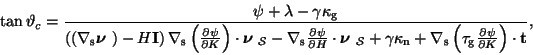
Figure 10. Sketch of a border.
Of course, the molecular splay has an energetic cost: according to Helfrich [13],
it can be given the simple form

Figure 11. An open fluid membrane that adheres to a rigid
wall.
![]() is the free membrane and
is the free membrane and
![]() the
adhering one;
the
adhering one;
![]() is the adhering contour, and
is the adhering contour, and
![]() the adhesive border of
the adhesive border of
![]() .
The principal curvatures of the membrane may
jump along
.
The principal curvatures of the membrane may
jump along
![]() .
The unit normal
.
The unit normal
![]() to
to
![]() is the same as the unit normal to the wall.
is the same as the unit normal to the wall.
On the other hand, an open membrane could also adhere to the wall along the border and be completely detached elsewhere (see Figure 12).
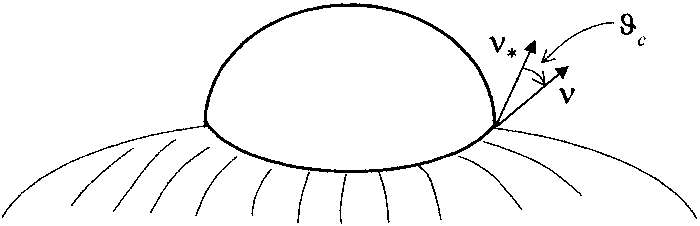
Figure 12. An open fluid membrane in contact with a rigid
wall only along its border
![]() .
Here, there is neither an adhering contour
nor an adhering membrane. The angle
.
Here, there is neither an adhering contour
nor an adhering membrane. The angle
![]() that the normal to
that the normal to
![]() makes with the normal to the wall along the border of the
membrane is the contact angle.
makes with the normal to the wall along the border of the
membrane is the contact angle.
In this case, the admissible displacements of the border can no longer be tangent to the membrane.
Starting from (1) and (2), one can derive a general equilibrium equation for a free membrane, which reduces to Ou-Yang and Helfrich's [14] when their special energy functional is used. This equation was obtained in [15] through an intrinsic method, that is, which does not resort to any specific system of coordinates to represent the equilibrium shape of the membrane:
In (18)
![]() ,
div s and
,
div s and
![]() denote,
respectively the surface Laplacian, the surface divergence, and the surface
gradient;
denote,
respectively the surface Laplacian, the surface divergence, and the surface
gradient;
![]() is the unit normal vector to the membrane and
is the unit normal vector to the membrane and ![]() is a
Lagrange multiplier which accounts for the inextensibility of the membrane.
Moreover,
is a
Lagrange multiplier which accounts for the inextensibility of the membrane.
Moreover,
![]() is the mean curvature and
is the mean curvature and
![]() he Gaussian curvature.
he Gaussian curvature.
Besides this equation, an open membrane in contact with a wall must also
satisfy other equilibrium conditions on both its possible adhering contours
and its adhesive borders. The former equilibrium condition reads as
On an adhesive border ![]() ,
when the membrane is everywhere tangent to the
wall (see Figure 11) the equilibrium condition reads as
,
when the membrane is everywhere tangent to the
wall (see Figure 11) the equilibrium condition reads as
When axisymmetric membranes are considered and ![]() is taken as
is taken as
As an application of (20), we have studied in [15] the stability for an adhesive border, when the adhesive body is a cone, as illustrated in Figures 13a & 13b
 b)
b)

Figure 13. The adhesion of a membrane to two axisymmetric
bodies: the cones in (a) and (b) are reverse to one another, but
with the same
amplitude ![]() .
The equilibrium of the border
.
The equilibrium of the border
![]() is unstable
for the former and stable for the latter.
is unstable
for the former and stable for the latter.
The analysis of (20) allows us to conclude that the equilibrium configuration shown in Figure 13a is unstable, whereas that shown in Figure 13b is stable.
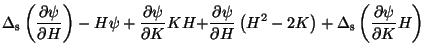
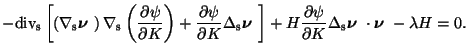
![$\displaystyle [\hspace{-0.05cm}[\psi]\hspace{-0.05cm}]\mbox{\boldmath$\nu$ }_{\...
...rtial K}\mathbf{I}]\hspace{-0.05cm}]\mbox{\boldmath$\nu$ }%
_{\mathcal{S}^{*}}-$](img86.png)